JOURNAL OF CRUSTACEAN BIOLOGY 19(4):738-751 (1999).
METHOD FOR ASSESSING RELATIVE ABUNDANCE, SIZE DISTRIBUTION, AND GROWTH OF RECENTLY SETTLED AND EARLY JUVENILE LOBSTERS (HOMARUS AMERICANUS) IN THE LOWER INTERTIDAL ZONE
Diane F. Cowan
ABSTRACT
The purpose of this study was to establish a method for repeated, year-round sampling of the abundance of young-of-the-year and juvenile lobsters, Homarus americanus, where they can be found intertidally. The primary advantage of the intertidal lobster monitoring program is the ability to overcome limitations pursuant to subtidal sampling techniques through increased temporal and spatial resolution of data collected, using sampling methods possible at low tide. Lobsters were sampled monthly throughout the years 1993-1997 by overturning rocks in one-square meter quadrats placed along fixed transects running parallel to the water's edge 0.2-0.4 m below mean low water (MLW). Individuals were tagged using binary-coded microwire tags. Two distinct size classes of lobsters, the smaller measuring 3-15 mm in carapace length (CL), the larger measuring 16-40 mm CL, were found consistently. Monthly mean densities ranged from 0-8.6 individuals per square meter at 0.4 m below MLW. These densities are comparable to published reports of similar-sized lobsters sampled at depths of 5 m below MLW. A seasonal pattern of abundance was observed, with the highest average densities recorded in May-November. Lower abundance in December-April was most likely due to seasonal migrations of individuals of the larger size class. Preliminary results indicate that areas of the lower intertidal zone serve as nursery grounds where postlarval lobsters settle and grow for several years. The assessment method developed in this study is being used to establish a time series that may be useful in developing and testing predictive models of annual yields and recruitment to the lobster fishery. The close proximity of intertidal lobster nurseries to the terrestrial zone indicates serious implications for fisheries management and issues of habitat protection in the shoreland zone. Further investigations are needed to yield a more complete picture of the full geographical extent of use of the intertidal zone as a nursery area for lobsters.
Introduction
The American lobster Homarus americanus is among the most economically valuable commercial fisheries resources along the northeastern coast of North America. For more than a century, efforts have been made to understand the factors that limit lobster abundance and thereby restrict recruitment to the fishery. Models have been developed and data gathered in an attempt to find reliable predictors of sustainable yield (reviewed in Fogarty, 1995; Bannister et al., 1996). The current working model proposed by Fogarty and Idoine (1986) summarizes the relationships between egg production, larval abundance, juvenile abundance, and recruitment to the fishery, and predicts that density-dependent factors regulating population size act on the early juvenile life-history stages. Suggested density-dependent factors include habitat limitations, predation, and competition.
In order to identify factors that may limit lobster abundance and restrict recruitment to the fishery, it is necessary to coordinate long-term studies (~10 years or more) that can provide the basis for time-series analyses. Other than landings records and trawl surveys conducted by the National Marine Fisheries Service, there exists a general lack of long-term studies that attempt to assess the American lobster population (Burns et al., 1983; NEFSC, 1996). In addition, there is a lack of coordinated effort in the collection, analysis, and interpretation of data gathered through recently begun short and medium-term studies (Estrella, unpublished; Hudon, 1987; Wahle and Steneck,1991; Watson and Howell, 1991; NEFSC, 1996; Wahle and Incze, 1997). Long-term studies are of particular importance to the study of the American lobster H. americanus due to the prolonged development time of an estimated seven years from benthic recruitment to sexually maturity.
Dye (1998) cited many benefits of establishing time series in terms of understanding the dynamics of intertidal communities. The benefits of long-term studies include providing a basis for: (1) interpreting the results of short-term studies, (2) detecting important processes that take place over long periods of time, such as the life cycle of the lobster, (3) understanding the impact of unusual events, and (4) investigating nonlinear phenomena such as chaos. In addition to improving the temporal context provided by long-term studies, there is a need for increasing the spatial scale to cover a broader geographical area. The issue of temporal and spatial scaling for assessment of intertidal habitat has been discussed elsewhere (McCook and Chapman, 1993).
In spite of the identification of postlarvae as the critical stage for lobster recruitment, measures of abundance (reported as mean number of individuals per m2) of the earliest juvenile stages (< 10 mm CL) have appeared only over the past decade (Hudon, 1987; Able et al., 1988; Heck et al., 1989; Wahle and Steneck, 1991). Various methods have been used to quantify benthic abundance, including visual surveys and airlift suction sampling (reviewed in Lawton and Lavalli, 1995).
The purpose of this paper is to introduce a new assessment tool and to evaluate its usefulness based on comparisons with preexisting methods for determining the abundance and distribution of recently settled and early juvenile H. americanus. Sampling in the lower intertidal zone provides data with increased temporal and spatial resolution. Previous studies have focused on sampling during the summer months, resulting in a general inability to detect seasonality of benthic recruitment, migrations, and other critical behavioral events that are essential to a complete understanding of the ecology of an organism. This study presents a methodology that allows year-round sampling that quantitatively measures the seasonal abundance of juvenile lobsters, H. americanus, from the time of settlement to approximately age five. The intertidal lobster monitoring program was designed to develop a method to measure the abundance of young-of-the-year and follow growth and small scale movements of subsequent juvenile lobsters in order to: (1) identify significant lobster nursery areas for habitat protection, (2) advance our knowledge of basic ecology and behavior of juveniles, (3) develop a prolonged time series that may be useful in forecasting future landings, and (4) provide quantitative data that may provide the basis for better informed management decisions.
MATERIALS AND METHODS
The discovery that lobsters use intertidal nursery habitats has led to the design of a sampling program (intertidal lobster monitoring program) that can augment subtidal sampling to increase the temporal and spatial resolution of data collection. SCUBA and snorkeling studies present several limitations, especially in the cold, turbid waters of the Gulf of Maine where year-round sampling is often prohibitory.
Regional Surveys.—Three surveys were conducted to identify the location of intertidal areas inhabited by young-of-the-year and juvenile lobsters (Figs. 1-3). Two of the surveys focused on determining the geographical extent of use of the intertidal zone as lobster nurseries, and the third was a long-term monitoring study. The geographical survey sites were similar to the long-term monitoring site in terms of habitat type, slope, degree of exposure to open water, accessibility to observers, and proximity to adults as evidenced by the presence of lobster trap gear in adjacent subtidal areas. Sampling took place during spring low tides and was accomplished by overturning rocks exposed along the low-tide terrace. Each lobster captured was measured in situ, identifying characteristics were recorded, and lobsters were immediately returned to their rock shelters.
The first survey covered areas of the Northwest Atlantic coastline from Long Island Sound to the Bay of Fundy. Sites throughout New England were surveyed between 20 July and 16 October 1993. Effort at each survey site was restricted to 2 h of sampling by 1 - 2 observers.
The second survey determined whether a predictable pattern of distribution could be ascertained on a more localized level. A pattern was sought as the basis for comparison with other sites and to provide the ability to make predictions about where other nurseries would occur throughout the Gulf of Maine. Sites concentrated midway through the range (along the coastline of the Town of Harpswell, Maine, in Casco Bay) were surveyed between 11 September and 5 November 1995. Survey sites varied according to compass heading to test the prediction that wind advection and surface currents may influence settlement (Harding et al, 1983; Harding and Trites, 1987; Wahle and Incze, 1997). Effort at each site was restricted to 2 h of sampling by 2 - 4 observers.
Long Term Study.—The long-term monitoring site was surveyed by sampling 1-m^2 quadrats along fixed horizontal transects during the 14 weeks of the lowest tides of each year (i.e., during all tides that were predicted to fall a minimum of 0.2 m below mean low water). Quadrat sampling began approximately 1 h before the predicted time of low water and continued for at least 1 h thereafter. Flash lights and headlamps were used to accomplish nocturnal surveys. This intensity of sampling would be prohibitive using snorkeling or diving techniques due to low water temperatures much of the year, poor visibility, time constraints, and agility of lobsters in the subtidal zone.
Lowell's Cove, on Orr's Island in Casco Bay, Maine, is a south-facing, shallow-water cove with a gently sloping intertidal zone experiencing vertical tidal fluctuations of approximately 3 m and an extreme tidal range of 3.6 m (Fig. 3). The cove is partitioned into two smaller inlets by a bedrock groin (Barleyfield Point in Fig. 3). The low-tide terrace sampled consisted of approximately 80% rocks with scattered pebbles on a substrate of silt to fine sand. The sampling area at transect 1 (easternmost transect in Fig. 3), where the highest densities of lobsters were found, consists of an island of loose pieces of flat, algae-covered bedrock between mudflat higher in the intertidal zone and silt with patches of Zostera marina L. in the shallow subtidal zone.
Monthly quadrat sampling along a minimum of 4 of 7 transects (numbered 1-4 from east to west in Fig. 3) was conducted when tides ranged from -0.2 to -0.6 m. The shallow subtidal zone was considered to start at -0.6 m, where the Chondrus zone abruptly ends (sensu Lubchenco and Menge, 1978). The area sampled each month varied with lobster abundance, due to time constraints of measuring and tagging individuals in the field. In the winter months, when lobster encounters were rare, all 7 transects (~2001 m^2 quadrats) could be sampled in a single set of spring tides. In contrast, in the summer months, as few as 50 quadrats along 4 transects were completed each month.
Numbered from east to west, transects 1 and 4 extended approximately 40 m along the water's edge, running east to west. Transects 2 and 3 extended approximately 20 m southward along east and west sides of the bedrock groin (Barleyfield Point, Fig. 3). Air, water, and substrate temperatures were recorded to the nearest 0.1°C using a Hannah instruments H19063 microcomputer K-thermocoupled thermometer at the onset and end of each sampling period. Surface salinity of the cove was measured at the same time, using a temperature-compensated handheld refractometer (American Optical Corporation). Whenever possible, salinities and temperatures were also measured in lobster burrows.
A qualitative description of each quadrat was made by estimating the percentage of rock cover and substrate type, the presence of each macroalgal and animal species observed, and the presence or absence of depressions filled with water. Movable rocks in the quadrats were then overturned one at a time and organisms found beneath the rocks were added to the species list.
When juvenile lobsters were found on the surface of dry sediments or in pools of water beneath the rocks, the following information was recorded: carapace length - (from the rear of the eye socket to the posterior margin of the carapace), total length (from the tip of the rostrum to the tip of the telson), handedness (right or left crusher), sex (for lobsters measuring >15 mm carapace length), condition of appendages (i.e., missing, regenerating, damaged), molt condition (appearance of exoskeleton and limb buds; Aiken, 1973; Waddy et al, 1995), condition of tissues (i.e., evidence of swelling at low salinities), and activity (any movement of appendages or entire body). Carapace length was measured to the nearest 0.5 mm using vernier calipers and total length to the nearest 1 mm using a hand ruler. Starting in September 1993, lobsters measuring at least 12 mm CL were tagged in situ by implanting an individually coded microwire tag (Northwest Marine Technology, Shaw Island, Washington), into the muscle tissue in the distal segment of the second right pereiopod (Krouse and Nutting, 1990). Each lobster was immediately returned to its shelter. In addition to quadrat sampling, lobsters were gathered through haphazard sampling higher in the intertidal zone, in tide pools, and in other regions of the cove in an attempt to recapture tagged lobsters. Lobsters could not be tagged when the air temperature fell below 5°C due to their propensity for autotomizing limbs at cold temperatures. All data were recorded on Sony Microcassette™ 3MC-60 audio cassette tapes with recording speed set for a capacity of 120 min. Tapes were later transcribed onto data sheets and the information was then entered onto a Microsoft Excel spreadsheet. Recapture analyses were done using SYSTAT.
Estimates of densities in number/m were based on quadrat sampling along all transects. However, for comparison to subtidal sampling, only transect l was used, since it had the greatest uniformity in substrate type surveyed subtidally elsewhere. Table 1 shows comparisons of densities are based on data collected in July, August, and September to correspond to subtidal studies that focused on the summer months. In Fig. 6, however, densities are based on data collected in September, October, and November, because this is the time of year during which settlement was typically observed (see Fig. 5).

Fig. 1. Distribution map of juvenile lobsters, Homarus americanus, found in the lower intertidal zone along the coast of New England. Juvenile lobsters were found living at the low intertidal shallow subtidal interface from Long Island Sound, Connecticut, to Winter Harbor, Maine. Filled circles indicate areas where juvenile lobsters were found. Open circles indicate areas where juvenile lobsters were not found. Dashed line indicates US/Canadian border. Area surrounded by the box indicates Harpswell, Maine. Original GIS map provided by the National Marine Fisheries Service.
RESULTS
Distribution of Juvenile Lobsters in the Intertidal Zone
The lobster Homarus americanus inhabited the low intertidal-shallow subtidal interface from Long Island Sound, Connecticut, to Winter Harbor, Maine (Fig. 1). Midway along this range, in Harpswell, Maine, a localized distribution pattern was discerned, with juvenile lobsters being found at intertidal sites exposed to the open waters of Casco Bay, but not farther up the inner bays (Fig. 2). The distribution pattern observed is most likely due to a combination of biological and physical factors affecting the settlement of postlarvae and the behavior of first-year juveniles (see Discussion).
Community Structure of Juvenile Lobster
At the long-term study site (Fig. 3), two modal size-class distributions were found consistently (Fig. 4). The smaller size class of individuals, measuring between 4 and 15 mm CL, represent young-of-the-year. Young-of-the-year juveniles appear to overwinter in the lower intertidal zone and can be found year-round (Fig. 5). Larger juveniles, measuring between 16 and 40 mm CL, were common from May through November, but rare from December through April. Larger subadult lobsters were occasionally found in small numbers (Fig. 4).
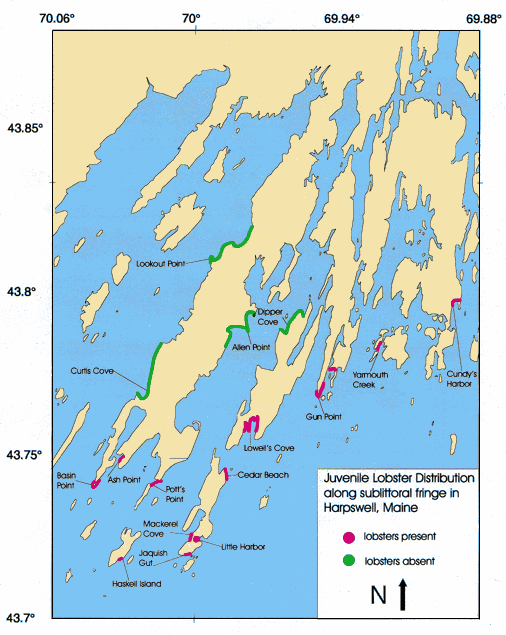
Fig. 2. Distribution map of juvenile lobsters, Homarus americanus, found in the lower intertidal zone along the coast of Casco Bay in Harpswell, Maine. Outlined borders indicate survey areas where lobsters were (filled border) and were not (open border) found. Original GIS map provided by the National Marine Fisheries Service.
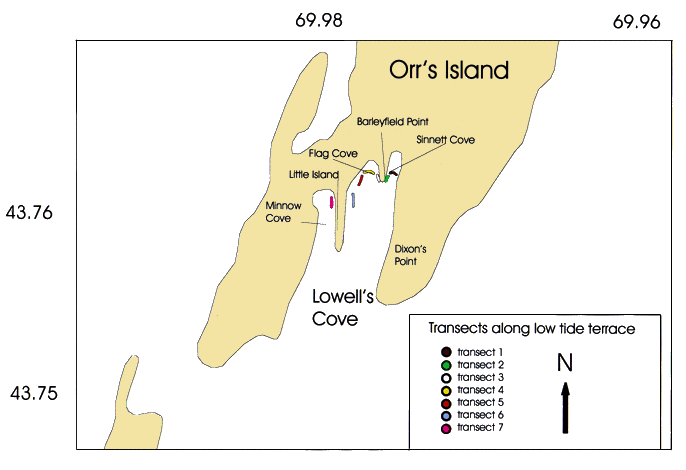
Fig. 3. Map of long-term monitoring site at Lowell's Cove on Orr's Island, Maine, indicating location of transects (numbered from east to west in text) and major geographic features where juvenile American lobsters, Homarus americanus, were sampled monthly from 1993-1997.
The abundance of lobsters of all size classes increased from 1993-1995 (Fig. 6). Overall densities and densities of young-of the-year (YOY) peaked in 1995, the warmest year since sampling began. The decrease in yearling density in 1996 was followed by a decrease in the larger size class with a one-year time lag. A further decrease in YOY density was observed in 1997 (Fig. 6).
The sex ratio was skewed toward males (X^2 goodness of fit; P > 0.001) at Lowell's Cove with 53% male, 30% female, and 17% of undetermined sex (Fig. 7). Sex is not determined until lobsters are several months old. Therefore, the youngest individuals sampled are accounted for in the 17% category. The incidence of injury is relatively low for intertidal lobsters, as evidenced by most lobsters having both claws (Fig. 8).
The average density at transect 1 for all years and all months combined was approximately 2 indiv / m^2 (Fig. 9). Monthly mean densities were generally higher than the overall mean during May through November. Densities fell below the annual mean during the winter months (December through April). This observation is explained by the sudden and abrupt disappearance of migratory juveniles during November. The absence of migratory juveniles in the winter is most likely due to movements into subtidal areas.
Tag and Recapture
Of the 6,872 lobsters sampled, 5,533 lobsters were individually tagged using binary coded microwire tags. Of the 5,533 marked lobsters, 434 were recaptured, yielding an overall recapture rate of 0.078 or approximately 8%. Many of the recaptured lobsters had molted at least once since the time of first capture, as evidenced by incremental changes in length (Fig. 10). The data in Fig. 10 show that the change in carapace length tends to be greater for larger individuals. The high degree of variability in the data is largely due to differences in the time interval between first and subsequent capture.
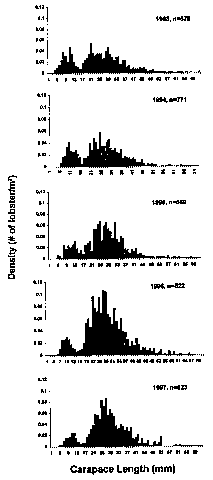
Fig. 4. Densities (number/m^2) plotted as size-frequency distribution of the lobster Homarus americanus captured in the lower intertidal zone at Lowell's Cove on Orr's Island, Maine, from 1993-1997. Carapace length plotted on x-axis extends to size of largest individual measured. N (n) = number of 1 m2 quadrats sampled.
|
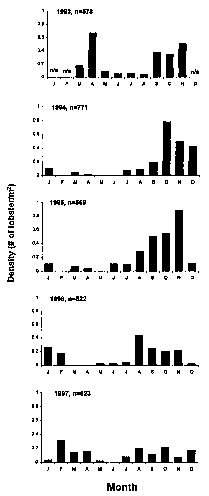
Fig. 5. Time of occurrence of young-of-the-year lobsters, Homarus americanus, indicated by plotting density (number/m^2) of lobsters measuring s15 mm CL found each month. N (n) = number of 1 m^2 quadrats sampled. na = data not available.
|
The time interval between first and second capture varied from 30-900 days and the change in carapace length varied from 0-22 mm over the same period of time (Fig. 11). As expected, change in carapace length increased with the number of days at large. The greatest changes in carapace length were recorded for individuals who escaped recapture for the longest duration. Although most individuals were captured only once or twice, six lobsters were captured on multiple occasions (Fig. 12). For example, female lobster number 2 (Fig. 12) was captured on five separate occasions. At the time of initial capture on 23 July 1994, lobster number 2 measured 18 mm CL. She was next captured on 14 June 1995 at a new size of 23 mm CL. During the summer of 1995 female number 2 underwent two additional molts, growing to 27.5 mm CL by 12 July 1995 and 31 mm CL by 9 September 1995. Each time female number 2 was captured in 1994 and 1995, she was at transect 1 within 7 m of the point of initial capture. Female number 2 was not captured again until 6 June 1996, after 270 days at large when female number 2 was found approximately 50 m away at transect 2, but had not molted. Individuals 2, 4, and 6 were each captured at 30-day intervals in the summer months. Their incremental changes in carapace length over that time period most likely represented the growth increase of one molting event. The long periods of time when numbers 1, 2, and 3 did not grow (in over 200 days at large) represent winter months. Overall, there appears to be a pattern for individuals of these sizes to molt two to three times per year, during the summer months (Fig. 12).
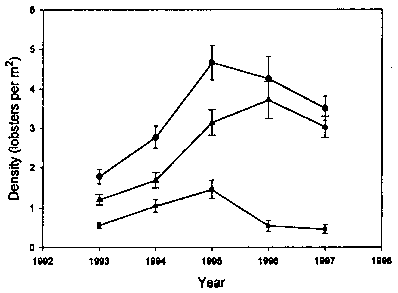
Fig. 6. Time series of densities (number/m: +- SE) of all lobsters (Homarus americanus) young-of-the-year (~ 15 mm CL), and larger juveniles (> 15 mm CL) found along transect 1 from September-November of each year, 1993-1997.
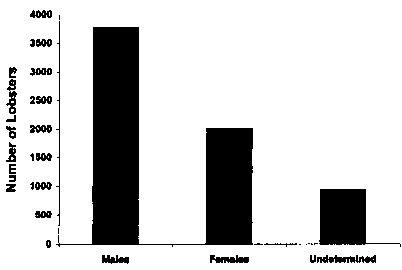
Fig. 7. Sex of juvenile lobsters, Homarus Americanus, at Lowell's Cove compiled over a five-year period from 1993-1997. Undetermined refers to individuals for which sex could not be determined due to body size and/or because sexual differentiation had not yet occurred. The sex ratio was skewed toward males (X goodness of fit, P > 0.001).
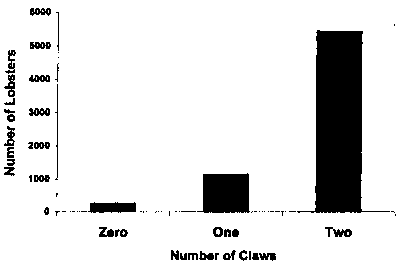
Fig. 8. Incidence of injury measured by recording claw loss for intertidal lobsters, Homarus americanus, at Lowell's Cove compiled over a five-year period from 1 993-1 997.
DISCUSSION
Significance of Method as an Assessment Tool
The results presented here introduce a new method for monitoring juvenile lobsters where they can be found intertidally and exemplify the power of using land-based survey methods to assess abundance, time of settlement, and growth of juvenile Homarus americanus for the first 4-5 years of life. The intertidal lobster monitoring program (ILMP) takes advantage of the use by lobsters of the lower intertidal zone in order to implement an intensive, long-term, spatially extensive survey of benthic recruitment and growth. The greatest advantage of the method is the ability to sample at any time of year for long sampling periods regardless of most environmental conditions, including photoperiod and foul weather. Inclement weather can restrict other sampling methods, including diving, trawl surveys, and trap studies. Intertidal sampling also allows for sampling with minimal disturbance to both the lobsters and their habitat. Using this method, lobsters are handcaptured with ease, tagged in situ, and immediately returned to their shelters. Although the ILMP may be useful as a powerful assessment tool, it is prudent to keep in mind that the intertidal zone represents the most landward margin of lobster distribution and thus may not be representative of the entire population. Therefore, the intertidal lobster monitoring program should be used in conjunction with subtidal studies.
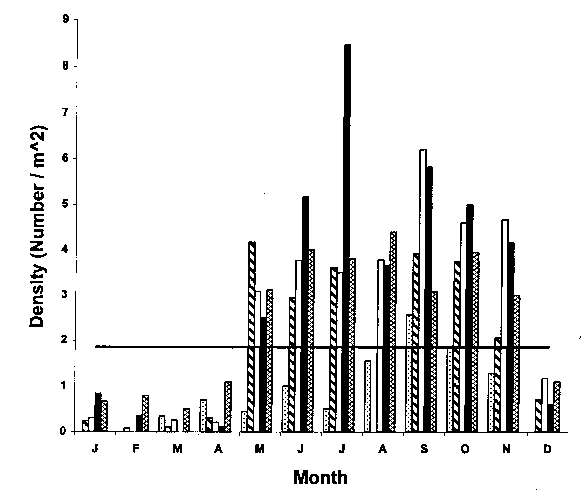
Fig. 9. Mean lobster density (number / m^2) at transect 1 for intertidal lobsters, Homarus americanus, at Lowell's Cove over a five-year period from 1993-1997. Horizontal line through the graph represents average density for all five years combined. Dotted bars = 1993; diagonal bars = 1994; open bars = 1995; filled bars = 1996; cross-hatched bars = 1997.
Spatial Resolution of Intertidal Lobster Monitoring Program
In terms of spatial resolution or geographic extent of monitoring, intertidal sampling allows for rigorous, quantitative sampling methods spanning those areas of the coastline yielding the highest annual landings. Adult Homarus americanus range from the Straight of Belle Isle, Newfoundland, to offshore areas of North Carolina (Cooper and Uzmann, 1980). The highest landings occur in the Gulf of Maine, Nova Scotia, and the Gulf of St. Lawrence (Miller, 1995). During the initial 1993 survey, lobsters were found inhabiting the lower intertidal zone at 12 sites in five New England states: Maine, New Hampshire, Massachusetts, Rhode Island, and Connecticut (Fig. 1). The northeasternmost site (Winter Harbor, Maine) represents the farthest extent for young-of-the-year lobsters documented thus far in the United States. However, use of the lower intertidal zone has been previously documented at St. Mary's Bay in Nova Scotia (MacKay, 1926). Further investigations are underway to yield a more complete picture of the full geographical extent of use of the intertidal zone as a nursery area for lobsters. During 1996, community volunteers began to use this method at sites in Harpswell, Maine. Since then, the intertidal lobster monitoring program has expanded to more than 20 sites along the coastline from Odiorne State Park in New Hampshire to sites at Stonington, Deer Isle, Maine (Cowan, unpublished).
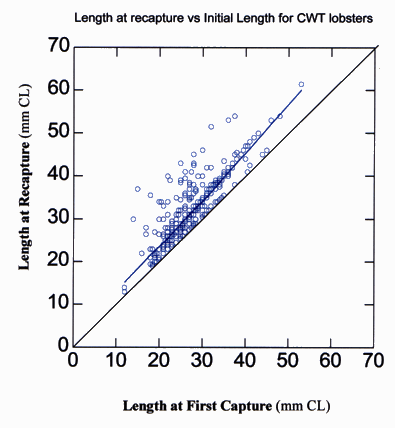
Fig. 10. Change in carapace length (mm CL) plotted as CL at first capture versus CL at subsequent capture of the lobster Homarus americanus tagged and recaptured using coded wire tags between September 1993 and December 1997. Each open circle represents one individual. Line drawn through circles indicates best fit for data. Line drawn through origin indicates the line of zero growth.
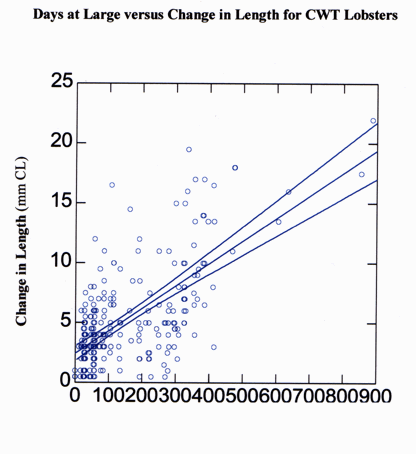
Fig. 11. Change in carapace length (mm) over time in number of days for the lobster Homarus americanus marked using coded wire tags between September 1993 and December 1997. Lines indicate best fit and 95% confidence interval.
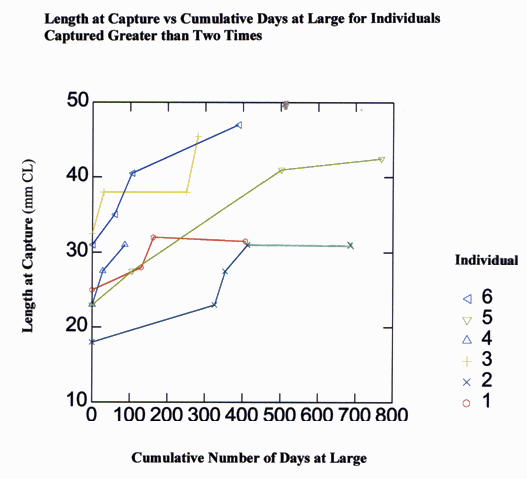
Fig. 12. Case histories of six individual juvenile lobsters, Homarus americanus, that were captured greater than two times showing change in carapace length over time relative to number of days at large. Open circles = lobster number 1; open triangles = lobster number 2; open squares = lobster number 3; filled circles = lobster number 4; filled triangles = lobster number 5; filled squares = lobster number 6.
In an effort to predict habitat requirements of intertidal lobsters, the survey of Harpswell, Maine, in 1995, uncovered juveniles at eleven out of fifteen sites (Fig. 2). Juvenile lobsters were found at sites exposed to the open waters of Casco Bay, but not along the shore line of inner bays and sounds (although marketable lobsters are fished adjacent to all sites surveyed). This distribution pattern suggests that biotic and abiotic factors may influence the suitability of locations as sites for benthic recruitment. Such factors may ultimately optimize growth rates and improve survivorship. Biological factors influencing suitability of settlement habitat may include proximity to larval supply, shelter availability as refuge from predators, and food availability. Potentially important physical factors include salinity, temperature, substrate type, wave action, ice scouring, and prevailing wind and water currents. Physical characteristics of settling sites exhibiting good intertidal habitat characteristics further upstream (i.e., inner bays) may be unsuitable as lobster nurseries due to extreme ice scouring, propensity of fresh-water runoff, and the amount of cover available in adjacent subtidal areas (mud bottom).
Temporal Resolution of Intertidal Lobster Monitoring Program
Observations of settlement, high densities, and multiple captures over the course of up to three years indicate prolonged use of the lower intertidal zone by individuals of H. americanus (Figs. 5, 9, 12). Most ecological observations of juvenile lobsters in the subtidal zone have been restricted to summer months (Hudon, 1987; Able et al., 1988; Heck et al., 1989; Wahle and Incze, 1997; Wahle and Steneck, 1991). Intertidal sampling allows for observations of daily and seasonal patterns of behavior. The densities of postlarvae and young-of-the-year (YOY defined as 3-15 mm CL, this study; see Lang et al., 1977; Hudon, 1987; Lavalli, 1991; Lavalli and Lawton, 1996; Palma et al., 1998, for other terminology used to describe this size class) and older juveniles (16-40 mm CL) varied over time. The relative proportion of older juveniles versus young-of-the-year increased in 1995 and 1996 (Fig. 9). Mean densities along transect 1 ranged from 0-8.6 indiv / m^2 (Fig. 8). A seasonal pattern of abundance was observed, with the highest densities being recorded from May through November. Lower densities in December through April were most likely due to seasonal migrations of yearlings and older juveniles to deeper water, since these individuals disappeared and reappeared abruptly. Further evidence for such movements is provided by results of the tagging study. For example, lobster number 2 (Fig. 11) was captured during three consecutive summer months, disappeared in autumn, and was never captured in winter. Other multiple recaptures were also not typically captured in winter. Further analyses of these recaptured lobsters may provide important information on growth rates, survivorship of year classes, and patterns of movements in terms of distances traveled over time.
Due to the ease of sampling out of the water, the ILMP method for measuring lobster abundance permits monthly resamples and thus results in an accurate time series that may be used to develop and test predictive models for annual yields and recruitment. An accurate time series can result because the sampling method is nondestructive; it does not disturb the sediments or remove organisms from the study site. Multiple resamples can also be made via suction sampling, but it takes a long time for the area sampled to recover, because the air lift removes the infaunal organisms as well as lobsters and a good part of the substrate. With repeated samples, the suction-sampled environment may become so degraded that it may resemble a desert patch where it is unlikely that postlarval lobsters will successfully settle. Therefore, in terms of developing a time series, subtidal suction sampling yields only a "scan" sample at a particular time. It is then necessary either to wait a long time, or to sample elsewhere. Sampling by overturning rocks in the lower intertidal zone gives on-going samples, month after month, year after year, in the same location.
Another shortcoming of subtidal sampling for benthic mobile organisms such as lobsters is the disturbance made during placement of sampling gear. Researchers monitoring lobster abundance subtidally "toss" a 0.5 m^2 quadrat and proceed with air-lift suction sampling (Wahle and Steneck, 1991; Palma et al., 1998). Mobile organisms on the surface of sediments will likely flee in response to such a disturbance. Although intertidal lobsters occasionally escape capture, the tail-flip escape response is far less useful out of the water.
Table 1. Examples of densities (+- 1 SD except where noted as SE) of lobsters (Homarus americanus) sampled in rocky substrates at shallow, coastal regions in Maine. In an attempt to compare similar substrate types within the same season, densities from this study were taken from transect 1 in July, August, and September.
Density (number/m^2)
5-40 mm CL |
Number of
quadrats |
Size of
quadrats |
Depth
(m below MLW) |
Location of Sample |
Year of
Sample |
Reference |
| 6.1 +- 5.0 SD |
23 |
0.25 m^2 |
5 |
Damariscove Island, Maine |
1987 |
Wahle and Stenek,1991 |
| 2.3 +- 2.8 SD |
24 |
|
10 |
|
|
|
| 6.9 +- 5.5 SD |
15 |
|
5 |
|
1988 |
|
| 3.0 +- 3.1 SD |
16 |
|
10 |
|
|
|
| 3.4 +- 4.0 SD |
20 |
0.25 m^2 |
5 |
Ocean Point, Maine |
1987 |
Wahle and Stenek,1991 |
| 1.6 +- 2.6 SD |
23 |
|
10 |
|
|
|
| 3.0 +- 3.4 SD |
16 |
|
5 |
|
1988 |
|
| 6.7 +- 4.5 SD |
16 |
|
10 |
|
|
|
| 0.1 +- 0.7 SD |
31 |
0.25 m^2 |
5 |
Pemaquid Point, Maine |
1987 |
Wahle and Stenek,1991 |
| 0.7 +- 1.6 SD |
23 |
|
10 |
|
|
|
| 1.5 +- 2.9 SD |
16 |
|
5 |
|
1988 |
|
| 1.5 +- 2.0 SD |
16 |
|
10 |
|
|
|
| 4.9 +- 0.7 SE |
12 |
0.46 m^2 |
3 - 6 |
Johns Bay and Damariscove Island, Maine |
1989 |
Incze and Wahle,1991 |
| 4.2 +- 0.9 SE |
12 |
|
|
|
1990 |
|
| 2.1 +- 2.2 SD |
61 |
1 m^2 |
0.4 |
Lowell's Cove, Orr's Island, Maine |
1993 |
Cowan, this study |
| 3.7 +- 2.0 SD |
25 |
|
|
|
1994 |
|
| 3.4 +- 2.3 SD |
49 |
|
|
|
1995 |
|
| 4.9 +- 2.5 SD |
46 |
|
|
|
1996 |
|
| 3.4 +- 1.9 SD |
36 |
|
|
|
1997 |
|
Significance of Lobster Use of the Intertidal Zone
Throughout its range, cover is a critical requirement for shelter-dependent juvenile lobsters that find refuge in various substrates, including cobble, rock, and boulder (Bernstein and Campbell, 1983; Hudon, 1987; Barshaw and Bryant-Rich, 1988; Barshaw and Lavalli, 1988; Wahle and Steneck, 1991, 1992), Zostera marina (see Barshaw and Lavalli, 1988; Heck et al., 1988; and personal observation), mud (Barshaw and Bryant-Rich, 1988; personal observation), and saltmarsh peat reefs (Able et al., 1988; Barshaw et al., 1994). Suitability of cover may vary with depth. For example, cobble beds are suitable for subtidal settling, but would not be useful intertidally, since small round rocks tend to roll in response to disturbances such as wave action (sensu Sousa, 1984). Intertidally, preferred rocks measured approximately 30 x 30 x 10 cm in dimensions and substrates found under lobster rocks included silt, mud, sand, and shell hash (Cowan, unpublished). Smaller sediment sizes have greater water retention and may therefore be preferred. However, many of the lobsters dug depressions where water pooled under their rocks, thereby increasing moisture.
Lobsters are likely to be found in intertidal areas where current speeds, salinities, and summer water temperatures are relatively high. Lobster nursery areas may occur as a band from the lower intertidal to shallow depths. In general, densities appear to be higher at depths of 0.4 and 5 m below mean low water than at depths of 10 m (Table 1). However, any such comparisons should be made with caution because the methods of data collection are not comparable temporally (sampling varies with season) and spatially (in terms of both geographical location and quadrat size). Comparisons have been attempted in the subtidal zone adjacent to the intertidal sampling site where hand-caught lobsters tended to be larger in size (Cowan, personal observation). However, smaller individuals may have escaped detection due to the different methods lobsters use for sheltering in various substrates. SCUBA diving during daylight hours revealed that lobsters in the adjacent subtidal zone excavated U-shaped burrows in the silt/mud substrate and were abundant in eelgrass beds (Cowan, personal observation).
Regardless of sampling method, postlarvae may not be expected to settle in deep water, due to the increased distance postlarvae would have to swim from the surface to the bottom. Greater distances from the surface waters to benthic settlement habitats may result in a higher rate of encounter with fish predators (Boudreau et al., 1992; Barshaw and Rich, 1997). The lower intertidal zone may represent an area of enhanced survivorship by acting as a refuge from high-predation intensity. Low abundance of large predatory fishes, characteristic of coastal waters nearshore (Witman and Sebens, 1992), may also play a role in the success of benthic recruitment. Although there is some predation in the intertidal zone, it may be less intense than in the subtidal zone. Bird predators, such as black-backed and herring gulls (Larus, sp.), are restricted to feeding on lobsters at certain times of the day only if the tide is low and seas are calm. In contrast, fish predators can actively hunt throughout the tidal cycle depending on their diurnal versus nocturnal activity levels.
Settlement in shallow water may optimize growth rates due to superabundant food supplies and elevated temperatures. Shallow waters tend to be warmer and growth rates of lobsters increase proportionally with increased temperature (Waddy et al., 1995, for review). The recapture data gathered using this method may be useful in resolving age cohorts and comparing growth rates and molt increments at various temperatures in the wild.
Potential Basic and Applied Significance of Intertidal Lobster Assessments
In terms of basic ecology, the ILMP may provide a model study for understanding the processes that underlie local population dynamics of a long-lived, large-bodied, benthic invertebrate. A full understanding of population dynamics requires knowledge of the rates of birth (recruitment), death, immigration, and emigration. Caley et al. (1996) argue that what is needed to understand the role of recruitment in open populations are multifactorial experiments in which recruitment and postrecruitment processes are simultaneously manipulated. However, before such experiments are appropriate it is necessary to have an indication of baseline levels of settlement and abundances. The ILMP can provide such baseline data for the American lobster.
Taken together, the measures of abundance, observations of behavior, and tagging data collected using this method may lead to an accurate time series including numbers of individuals, resolution of age classes, and mortality estimates. Such information is critical in forecasting future stock abundance and detectable changes may serve as an early warning several years before stock collapse. Homarus is characterized as a long-lived genus, based on estimates of growth rates for H. americanus and field studies where hatchery-reared juveniles of H. gammarus (L.) were tagged at a few months of age, released into the wild, and subsequently recaptured in harvester's traps seven years later (Bannister et al., 1994). Based on the long time interval between benthic settlement and recruitment to the fishery, an impending stock collapse could go relatively unnoticed for several years in the absence of reliable predictors of sustainable yield.
Life history information is of particular value in the case of H. americanus due to the economic importance of the industry that the species supports throughout the northeastern coast of North America. The results of the intertidal lobster monitoring program indicate that effective protection of the lobster resource may rely not only on fishing regulations, but also on protecting coastal habitats. The intertidal zone is a particularly vulnerable habitat due to its close proximity to major anthropogenic impacts such as land sources of pollution, fresh water and road runoff, and erosion and sedimentation brought about through poor management practices in coastal development.
ACKNOWLEDGMENTS
I thank all of those who have offered advice and technical and financial support throughout the years of this study. Sara Kirby did many of the surveys in Maine and southern New England in partial fulfillment of the requirements for her Honor's Thesis at Bates College. Observers participating in the Harpswell surveys included community volunteers for the Harpswell Conservation Commission who were accompanied by DFC and recruited by Commissioner Josie Quintrell. Financial support for the volunteer program was provided by the Collaboration of Community Foundations for the Gulf of Maine, Commercial Fisheries Project, and the Davis Conservation Foundation. Special thanks go to Dr. Paul Rago at the Northeast Marine Fisheries Science Center in Woods Hole for assisting in compilation and analysis of the data; and to Lisa Hendrickson of NMFS for providing the GIS maps shown in Figures 1-3. I gratefully acknowledge the financial support of the Maine lobstermen who contribute to the Lobster Seed Fund of the Lobster Advisory Council of the Maine Department of Marine Resources.
LITERATURE CITED
Able, K.W., K. L. Jr. Heck, M. P. Fahay, and C. T. Roman. 1988. Use of salt-marsh peat reefs by small juvenile lobsters on Cape Cod, Massachusetts.—Estuaries 11: 83-86.
Aiken, D. E. 1973. Proecdysis, setal development and molt prediction in the American lobster (Homarus americanus).—Journal of the Fisheries Research Board of Canada 30: 1337-1344.
Bannister, R. C. A., J. T. Addison, and S. R. J. Lovewell. 1994. Growth, movement, recapture rate and survival of hatchery-reared lobsters (Homarus gammarus Linnaeus, 1758) released into the wild on the English east coast.—Crustaceana 67: 156-172.
—, L. W. Botsford, R. K. Mohn, V. R. Restrepo, and P. F. Sale. 1996. A review of the population dynamics of American lobster in the Northeast.—Atlantic States Marine Fisheries Commission Special Report 61: i-vi, 1-48.
Barshaw, D. E., and D. R. Bryant-Rich. 1988. A longterm study on the behavior and survival of early juvenile American lobster, Homarus americanus, in three naturalistic substrates: eel grass, mud, and rocks.—Fisheries Bulletin, United States 86: 789-796.
—, and K. L. Lavalli. 1988. Predation upon postlarval lobsters, Homarus americanus, by cunners, Tauogolabrus adspersus, and mud crabs, Neopanope sayi, on three different substrates: eelgrass, mud, and rocks.—Marine Ecology Progress Series 48: 119-123.
—, and D. R. Rich. 1997. An analysis of substrate selection by postlarval American lobster, Homarus americanus, using a dynamic optimization model.— Oikos 80: 554-564.
—, K. W. Able, and K. L. Heck. 1994. Salt marsh peal reefs as protection for postlarval lobsters, Homarus americanus, from fish and crab predators: comparisons with other substrates.—Marine Ecology Progress Series 106: 203-206.
Bernstein, B. B., and A. Campbell. 1983. Contribution to the development of methodology for sampling and tagging small juvenile lobsters (Homarus americanus).—Canadian Manuscript Report of Fisheries and Aquatic Sciences, Number 1741, pp. 1-34.
Boudreau, B., Y. Simard, and E. Bourget. 1992. Influence of a thermocline on vertical distribution and settlement of post-larvae of the American lobster Homarus americanus Milne-Edwards.—Journal of Experimental Marine Biology and Ecology 162: 35-49.
Burns, T. S., R. Schultz, and B. E. Brown. 1983. The commercial catch sampling programs for the American lobster in the U.S. Northwest Atlantic.—In: W. F. Doubleday and D. Rivard, eds., Sampling commercial catches of marine fish and invertebrates.—Canadian Special Publication Fisheries and Aquatic Sciences. Pp. 82-95
Caley, M. J., M. H. Carr, M. A. Hixon, T. P. Hughes, G. P. Jones, and B. A. Menges. 1996. Recruitment and the local dynamics of open marine populations.—Annual Review of Ecology and Systematics 27: 477-500.
Cooper, R. A., and J. R. Uzmann. 1980. Ecology of juvenile and adult lobsters, Homarus.—In: J. S. Cobb and B. E Phillips, eds., The biology and management of lobsters. Vol. 2, pp. 97-142. Academic Press, New York, New York.
Dye, A. H. 1998. Dynamics of rocky intertidal communities: analyses of long time series from South African shores.—Estuarine, Coastal and Shelf Science 46: 287-305.
Fogarty, M. J. 1995. Populations, fisheries, and management.—In: J. R. Factor, ed., Biology of the lobster, Homarus americanus. Pp. 111-151. Academic Press, New York, New York.
— and J. S. Idoine. 1986. Recruitment dynamics in an American lobster (Homarus americanus) population.—Canadian Journal of Fisheries and Aquatic Sciences 43: 2368-2376.
Harding, G. C., and R. W. Trites. 1987. Dispersal of Homarus americanus larvae in the Gulf of Maine from Browns Bank.—Canadian Journal of Fisheries and Aquatic Sciences 45: 416-425.
—, K. F. Drinkwater, and W. P. Vass. 1983. Factors influencing the size of American lobster ( Homarus americanus) stocks along the Atlantic coast, Nova Scotia, Gulf of St. Lawrence, and Gulf of Maine; a new synthesis.—Canadian Journal of Fisheries and Aquatic Sciences 40: 168-184.
Heck, Jr., K. L., K. W. Able, M. P. Fahay, and C. T. Roman. 1989. Fishes and decapod crustaceans of Cape Cod eelgrass meadows: species composition, seasonal abundance patterns and comparisons with unvegetated substrates.—Estuaries 12: 59-65.
Hudon, C. 1987. Ecology and growth of postlarval and juvenile lobster, Homarus americanus, off Iles de la Madeleine (Quebec).—Canadian Journal of Fisheries and Aquatic Sciences 44: 1855-1869.
Incze, L. S., and R. A. Wahle. 1991. Recruitment from pelagic to early benthic phase in lobsters, Homarus americanus.—Marine Ecology Progress Series 79: 77-87.
Krouse, J. S., and G. E. Nutting. 1990. Evaluation of coded microwire tags inserted in legs of small juvenile American lobsters.—American Fisheries Society Symposium 7: 304-310.
Lang, F., C. K. Govind, W. J. Costello, and S. 1. Greene. 1977. Developmental neuroethology: changes in escape and defensive behavior during growth of the lobster.—Science 197: 682-685.
Lavalli, K. L. 1991. Survival and growth of early juvenile American lobsters Homarus americanus through their first season while fed diets of mesoplankton, microplankton, and frozen brine shrimp.—Fishery Bulletin, United States 89: 61-68.
—, and P. Lawton. 1996. Historical review of lobster life history terminology and proposed modifications to current schemes.—Crustaceana 69: 594-609.
Lawton, P., and K. L. Lavalli. 1995. Postlarval, juvenile, and adult ecology.—/n: J. R. Factor, ed., Biology of the lobster, Homarus americanus. Pp. 47-88. Academic Press, New York, New York.
Lubchenco, J., and B. A. Menges. 1978. Community development and persistence in a low rocky intertidal zone.—Ecological Monographs 48: 67-94.
McCook, L. J., and A. R. O. Chapman. 1993. Community succession following massive icescour on a rocky intertidal shore: recruitment, competition and predation during early, primary succession.—Marine Biology 115: 565-575.
MacKay, D. A. 1926. Report on lobster investigations at St. Mary Bay, Digby County, N. S.—Biological Board of Canada. Pp. 1-6.
Miller, R. J. 1995. Fishery regulations and methods.— In: J. R. Factor, ed.,Biology of the lobster, Homarus americanus. Pp. 89-109. Academic Press, New York, New York.
North East Fisheries Science Center. 1996. Description of sampling programs for American lobster in the U.S. Northwest Atlantic.—NEFSC Stock Assessment Workshop Report 22: 1-56.
Palma, A. T., R. A. Wahle, and R. S. Steneck. 1998. Different early post-settlement strategies between American lobsters Homarus americanus and rock crabs Cancer irroratus in the Gulf of Maine.—Marine Ecology Progress Series 162: 215-225.
Sousa, W. P. 1984. The role of disturbance in natural communities.—Annual Review of Ecology and Systematics 15: 353-391.
Waddy, S. L., D. E. Aiken, and D. P. V. De Kleijn. 1995. Control of growth and reproduction.—In: J. R. Factor, ed., Biology of the lobster, Homarus americanus. Pp. 217-266. Academic Press, New York, New York.
Wahle, R. A., and L. Inzce. 1997. Pre- and post-settlement processes in recruitment of the American Lobster.—Journal of Experimental Marine Biology and Ecology 217: 179-207.
— and R. S. Steneck. 1991. Recruitment habitats and nursery grounds of the American lobster Homarus americanus: a demographic bottleneck?—Marine Ecology Progress Series 69: 231-243.
—, and . 1992. Habitat restrictions in early benthic life: experiments on habitat selection and in situ predation with the American lobster.—Journal of Experimental Marine Biology and Ecology 157: 91-114.
Watson, W. W., and W. H. Howell. 1991. Seasonal movements of lobsters in the Great Bay Estuary. National Shellfisheries Association Program and Abstracts of 83rd Annual Meeting. Pp. 1-301.
Witman, J. D., and K. P. Sebens. 1992. Regional variation in fish predation intensity: a historical perspective in the Gulf of Maine.—Oecologia 90: 305-315.
RECEIVED: 10 August 1998.
ACCEPTED: 29 January 1999.
Please address all correspondence to The Lobster Conservancy in Maine.
Address:
The Lobster Conservancy, P.O. Box 235, Friendship, Maine 04547, U.S.A. (email:dcowan@lobsters.org)
Marine Policy Center, Woods Hole Oceanographic Institution, Woods Hole, Massachusetts 02543, USA